Artrópodes Cambrianos
Estudando Lobopodas um grupo de pesquisadores
conseguiu estabelecer que genes Hox estavam presentes em animais muito antes da
explosão cambriana no registro fóssil e o
Aysheaia, especialmente pela sua simplicidade anatômica o mais
requisitado.O Aysheaia tinha corpo mole, segmentações simples e claramente esta
relacionada ao grupo dos artrópodes.

Aysheia pedunculata
Dentro dos
Artrópodes, ocorre uma existência de Hexapoda-Crustácea que se encaixa com as análises de dados moleculares
anteriores. Uma comparação de sequências de DNAr 18s entre uma espécie de
Tardígrada e um grande número de sequências de artrópodes, outros
protostomiados, deuteróstomas, e um platelminto indicam claramente que os
Tardígrada estão intimamente relacionados com a linhagem dos artrópodes. Resultados
que suportam a ideia de eles e os artrópodes formam um clado monofilético.

O Xisto de Burgess, no Canadá, contém cerca de 140 espécies de animais do Cambriano dispostos em mais de 10 filos. Tanto o Xisto de Burgess, quantos os sítios de Chengjiang (15 milhões de anos mais velho do que o Burgess) apresentam uma espetacular diversidade de artrópodes e Lobopodas. Eles não têm pernas articuladas, mas estão diretamente relacionados aos artrópodes, apresentando uma segmentação simples, repetidas em grande quantidade, fruto da atuação de genes hox. Os artrópodes do Cambriano contem muitos segmentos corporais e apêndices associados a eles .
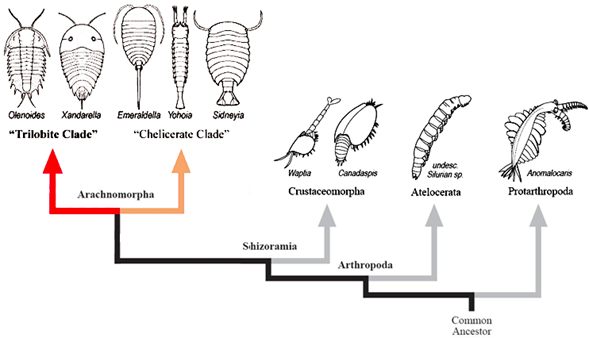
Os trilobitas e quelicerados tem um ancestral
comum com insetos miriápodos e crustáceos, obviamente trilobitas tem também 10
genes hox. Assim, trilobitas, quelicerados, miriápodes, crustáceos e insetos
formam um unido grupo, os artrópodes que tem um ancestral único, comum a todos
e que tem 10 genes Hox.
O animal que parece ser mais antigo que os artrópodes modernos e serviria como intermediário entre as Aysheaia e os artrópodes modernos seria
o onicóforo. Os onicóforos se
diversificaram durante o período de explosão de vida do Câmbrico. são um dos grupos mais primitivos e antigos
do que os artrópodes e poderiam servir como grupo irmão para uma análise
comparativa que poderia conectar os dois grupos a Aysheaia. Tanto onicóforos que são claramente
relacionados a Aysheaia e o Hallucigenia
quanto aos artrópodes modernos contém
também 10 genes Hox. Portanto é mais do que evidente que com essa característica eles descendam das Aysheaia, e que apesar de ser datados em 540 milhões de anos já possuíam os genes Hox
de seus ancestrais e a mais de 50 milhões de anos antes do inicio do Cambriano.
Yohoia


Yohoia é um gênero de animais
minúsculos e extintos do período cambriano que tem sido encontrado como fósseis
na formação de xisto de Burgess da Colúmbia Britânica, no Canadá. Foi colocado fileticamente entre os aracnomorfos, um grupo de artrópodes que inclui os quelicerados e possivelmente
os trilobitas, bem como aranhas e escorpiões modernos. Seus tamanhos variam de 7 a 23 mm. 711 espécimes de Yohoia são
conhecidos a partir do leito do Grande Fiópode,onde compreendem 1,35% da
comunidade.

Yohoia é um dos artrópodes do
"grande apêndice". Todos os espécimes possuíam um único par de grandes membros
articulados pré-orais com extremidades espinhosas ramificadas para agarrar,
empalar ou filtrar alimentos. Os Artrópodes com "grande apêndice" têm sido
vistos como um grupo polifilepético onde o apêndice foi desenvolvido independentemente,
ou como uma Megacheira de Classe incluindo Yohoia (com Leanchoilia,
Alalcomenaeus, Oestokerkus, Fortiforceps, Jianfengia, Yawunik e Isoxys)
definidos como euartrópodes, mais os anomalocarídeos, assim os definindo como um grupo
irmão aos artrópodes. Os gêneros semelhantes à yohoia são pequenos e têm
membros birramosos com um segmento ambulante, enquanto os anomalocarídeos são
animais maiores sem membros, exceto para o grande apêndice.


Estes animais tinham um "cotovelo" pronunciado e terminaram em quatro espinhas longas, parecendo bastante como dedos. Havia três apêndices na parte inferior do escudo da cabeça de cada lado, e supõe-se que eles apoiaram a criatura no fundo do mar arenoso ou silty (areia granulosa). Havia também apêndices únicos pendurados sob as placas do corpo que eram semelhantes a retalhos e franjas com setas, provavelmente usados para natação e respiração. Os espécimes também mostram algumas formações bulbosas na frente do escudo da cabeça que podem ter servido como olhos.
Acredita-se que Yohoia tenha sido uma
criatura principalmente bentônica (habitação no fundo do mar) que nadou logo
acima do fundo do oceano lamacento, usando seus apêndices para limpar ou
capturar presas.
Megacheira

Leanchoilia superlata. Haikoucaris
ercaiensis. Yohoia tenuis. Fortiforcepsos folia.
Megacheira é uma classe extinta de
artrópodes predatórios que possuía um par de grandes apêndices, daí o nome da
classe, bem como o nome comum "grandes artrópodes de apêndice". Suas
estruturas neurais e apêndices deutocerebral se assemelham aos de quelicerados.
A maioria deles foi encontrada em ambientes marinhos em todo o mundo, do
cambriano inferior ao médio.
A distinções entre megacheiran, grandes
apêndices e os apêndices cefálicos de outros artrópodes foram discutidos por
décadas. Há controvérsia sobre se eles são homólogos tanto para dinocaride
(radiodontes e lobopodianos gilled) e os apêndices frontais
do Isoxys (artrópode Bivalve) e dos quelicerados. Com base em evidências neuroanatômicas,
muitos estudos apoiam sua homologia para os quelicerados, e o surgimento das primeiras antenas de
outros artrópodes (que são deutocerebral),mas não os apêndices frontais
dinocarides (Um estudo de 2020 sugere que os grandes apêndices de
megacheiranos e radiodontes são homólogos e lança dúvidas sobre a validade de
suspeitas de evidências neuroanatômicas.

De cima a baixo Fortiforceps,
Sklerolibyone Jianfengia
A monofilia de Megacheira é incerta, e megacênbios são sugeridos como Quelicerados de grupos-tronco ou artrópodes de grupo-tronco, com a primeira hipótese baseada na morfologia semelhante as quelíceras e de grandes apêndices e ao lado da neuroanatomia e lábio reduzido o que os faz semelhante aos de um quelicerado moderno. Como exemplo da última hipótese, um estudo realizado em 2020 descobriu que a Megacheria formava um grupo de artrópodes mais diversificadoss do que dinocarídeos e isoxíferas, mas menos do que a coroa dos artrópodes, no basal que une quelicerados e miriápodes.
A A inclusão prévia de alguns gêneros "bivalves", como Forfexicaris, Ovalicephalus,e Occacaris a Megacheira foi questionada por investigações posteriores mas ainda aceita por falta de comprovações que os excluiriam do grupo.

Yawunik
kootenayi
Isoxycos


Os isoxycos (que significa "superfícies iguais") é um gênero de artrópode bivalve extinto pelágico; as várias espécies encontradas podem ter sido de predadores. Tinha um par de grandes olhos esféricos (que são a característica mais comumente preservada da anatomia de corpo mole), e dois grandes apêndices É possível que esses apêndices sejam homólogos dos grandes apêndices de radiodontes e megacheiras.

Isoxys acutangulus
Isoxys era abundante em mares
tropicais, e pode ter tido uma distribuição global. Olhos de diferentes espécimes parecem ter sido
adaptados a diferentes intensidades de luz; um espécime de Isoxys auritus era ou
crepuscular em águas rasas, ou vivia em águas cerca de 140 metros abaixo da
superfície do mar; enquanto outro foi morfologicamente adaptado a uma
intensidade de luz diurna em águas rasas. É provável que Isoxys era um predador visual
que caçava nadando acima do fundo do mar. Tinha poderosos apêndices frontais
preênseis e grandes olhos esféricos. O animal poderia impulsionar-se pela batida
possivelmente 14 pernas de natação em forma de nadadeiras e de se guiar com
uma cauda semelhante a um telson (peça quitinosa do exosqueleto).
Xandarella


O Xandarella spectaculum é uma espécie conhecida por muito poucos exemplos, alguns dos quais mostram muitos detalhes requintados. O espécime é preservado de forma dorsoventral, com as antenas e muitos apêndices bem preservados. O spectaculum é bastante trilobita na aparência, mas difere na falta de um exoesqueleto calcificado e por uma morfologia ocular diferente. O táxon é desconhecido fora do Biota de Chengjiang .


O animal pertence ao gênero extinto de
Xandarellid artiopodan conhecido no Cambriano da China e do Marrocos. Uma
espécie adicional Xandarella mauretanica foi descoberta em segmentos do
Cambriano no Marrocos preservado apenas a anatomia ventral. Como outros Xandarellids, o exoesqueleto é não
minado. O encéfalo pronunciou fendas oculares, presumivelmente derivadas de
olhos ancestrais perseguidos ventrais.

X.
mauretanica tipo espécime, mostrando apêndices biaramous
Sidneyia


Sidneyia é um
artrópode extinto conhecido por fósseis encontrados desde os xistos de Maotianshan do início do cambriano até a formação de xisto do médio Cambriano Burgess da Colúmbia
Britânica.

O Sidneyia variou de 51 a 127 mm de comprimento e é um dos maiores artrópodes encontrados no local. Acredita-se que tenha sido um carnívoro e coletor bentônico que caminhou pelo fundo do mar em busca de presas de casca dura. O conteúdo intestinal revelou que Sidneyia se alimentou em grande parte de pequenos trilobitas, bem como em braquiópodes, hiólitos (animais semelhante a molúsculos com concha) e pequenos artrópodes.

O intestino do Sidneya era estreito, mas aumentava posteriormente para formar uma bolsa, onde a digestão presumivelmente ocorria. A
retenção de fezes provavelmente indica alimentação pouco frequente Suas
gnathobases (estruturas para moer ou mandíbulas) requintadamente preservadas se assemelham às de Limulus, e
provavelmente foram usadas para esmagar presas.
Waptia fieldensis


Waptia fieldensis é uma espécie extinta de artrópode do Médio Cambriano da biota do Xisto de Burgess. Ele media cerca de 8 cm e se parecia com o camarão moderno, tanto na morfologia quanto no hábito a princípio. Com base no número de indivíduos, Waptia fieldensis é o terceiro artrópode mais abundante da Formação do Xisto de Burgess .
O animal possuia uma grande carapaça bivalvulada, um corpo segmentado terminando em um par de abas de cauda. Era um nadador ativo, alimentando-se de partículas orgânicas recolhidas do substrato do fundo do mar. É também um dos animais mais antigos com evidência direta de cuidado com a prole .
Canadaspis


Canadaspis ("Escudo do
Canadá") é um gênero extinto de crustáceo cambriano, um alimentador
bentônico aparentemente, que se movia principalmente caminhando e possivelmente usava seus
apêndices bi remos para agitar lama em busca de comida. O gênero foi colocado
dentro da subclasse Phyllocarida,na classe Malacostraca que inclui camarões e
lagostas atuais.


Espécimes de Canadaspis perfecta do Xisto
burgess
O Canadaspis tinha garras na
extremidade de seus apêndices frontais que podem ter sido usados para agitar
sedimentos, ou para raspar a camada superior, que pode ter sido uma camada
nutritiva de micróbios. Grandes partículas que ele agitava acabavam sendo capturadas por espinhos no interior de seus membros; essas espinhas teriam direcionado as
partículas de alimentos para a boca do organismo, onde usaria as suas mandíbulas
para moer partículas de maior tamanho

Suas antenas serviam a uma função
sensorial. Os espinhos em sua cabeça
provavelmente eram para proteger seus olhos, mais vulneráveis aos predadores. Seus
membros deveriam se mover em sequência para produzir um movimento
ondulado. Embora o Canadaspis provavelmente não nadava, isso poderia ter ajudado
a impulsionar o organismo sob sedimentos macios. Os apêndices também produziram
correntes que teriam ajudado na alimentação e respiração. O Canadaspis está muito
intimamente relacionado com o Perspicaris, diferindo apenas
em detalhes morfológicos.


Perspicaris é um artrópode fóssil do
período cambriano. Tinha 2-3 centímetros de comprimento e bivalved. As
válvulas, envolvendo o tórax,foram unidas por uma dobradiça dorsal.


Habelia Optata é um gênero de artrópode extinto do Cambriano Médio. Seus fósseis foram encontrados no Xisto de Burgess, na Colúmbia Britânica, Canadá.

O habelia media aproximadamente 02 cm de
comprimento com uma cauda, viveu durante o
período cambriano médio á cerca de 508 milhões de anos atrás. Como todos os
artrópodes, Habelia Optata apresenta um corpo segmentado com esqueleto externo
e membros articulados.
O Habelia Optata
possuia um corpo bem blindado, coberto por uma infinidade de espinhas
diferentes, sendo dividido em cabeça, tórax e pós-tórax, este todos com diferentes
tipos de apêndices. O tórax exibe cinco pares de pernas ambulantes, enquanto o
pós-tórax abriga apêndices arredondados provavelmente usados na respiração.
A cabeça de Habelia continha uma série
de cinco apêndices feitos de uma grande placa com dentes para mastigação, um
ramo semelhante a uma perna com espinhos rígidos como cerdas para agarrar, e um
ramo esguio e alongado modificado como um apêndice sensorial ou tátil.

Artrópodes que possuem estrutura desenvolvida para a
manipulação do alimento ao redor da boca, as quelíceras , chamam-se quelicerados, ao contrário dos insetos, crustáceos e
miriápodes, que, por possuírem mandíbulas, e são chamados de mandibulados. Apesar
da relação evolutiva de Habelia com os quelicerados, algumas características incomuns levaram pesquisadores a comparar a
cabeça de Habelia com a dos mandibulados no que se refere à uma perspectiva funcional.

Estrutura da cabeça do Habelia optata
Os ramos sensoriais peculiares podem
ter sido usados de forma semelhante à dos mandibulados, que usam antenas. Além
disso, os apêndices sobrepostos em forma de placa na série média de cinco são
mostrados para abrir e fechar paralelo à parte inferior da cabeça - assim como
eles fazem em mandibulados - especialmente aqueles que se alimentam de animais
com carapaças endurecidas.

Por fim, um sétimo par de apêndices na
parte de trás da cabeça parece ter cumprido uma função como os apêndices em
mandibulados, que auxiliam com os outros membros da cabeça no processamento de
alimentos. Essa ampla correspondência em função e não na origem evolutiva é
chamada de convergência, sendo assim do ponto de vista evolutivo, Habelia está
perto do ponto de divergência entre quelicerados e mandibulados.
Emeraldella

Emeraldella é um gênero de artrópode
conhecido do Cambriano Médio da América do Norte. A espécie tipo Emeraldella Brocki foi
descrita em 1912 do Xisto de Burgess one cerca de 21 espécimes deste animal são
conhecidos. Uma segunda espécie, o Emeraldella brutoni, fora encontrada no Xisto Wheeler, e que foi descrito em
2011. Um espécime adicional de E.
brutoni fora relatado em 2019, o que revelou mais da anatomia geral destes animais.
Emeraldella Brocki


Emeraldella Brocki atingia até 60 mm de
comprimento, possuia 12 tergites na seção do lombo, com um telson estendido. As antenas eram longas e podiam atingir o comprimento do corpo, além de serem compostas por
mais de 80 segmentos. Estruturas
oculares parecem estar ausentes nesta espécie.
Emeraldella Brutoni


O Emeraldella brutoni atingia até 50 mm de
comprimento, possuia cerca de 10 tergites de tronco, e possui Ocelos visíveis, ao contrário
de Emeraldella Brocki, as antenas não são preservadas bem o suficiente para dizer o
verdadeiro comprimento.
Leanchoilia


Leanchoilia é um artrópode Megacheirano ( tinham o sistema nervoso central muito parecido com os caranguejos-ferradura e os escorpiões atuais )conhecido nos depósitos cambrianos do Xisto de Burgess no Canadá
e da biota de Chengjiang da China. Media
cerca de 5 centímetros de comprimento e tinha longas antenas parecidas com chicotes montados em apêndices
frontais semelhantes a braços. Seus órgãos internos são ocasionalmente
preservados dentro do substrato em três dimensões.

Leanchoilia Superlata,
Leanchoilia Obesa
Leanchoilia Perséfone.
Eles podem, no entanto, ser exemplos de dimorfismo sexual. Há cerca de 55 espécimes de Leanchoilia conhecidos principalmente do Xisto de Burgess.

Marella


Marrella é um
gênero de artrópode encontrado por Charles Doolittle Walcott no Folhelho
Burgess. É o fóssil mais abundante do Folhelho. Apelidado erroneamente de
"caranguejo rendado" por seu descobridor, Marrella foi um artrópode
marinho que viveu no período cambriano.


Este organismo
possuía dois pares de antenas pré-orais e uma carapaça na cabeça com dois
espinhos voltados para trás. Após a cabeça, encontravam-se 24 a 26 segmentos
corporais birrêmes terminando em um minúsculo télson na extremidade posterior.
Os apêndices do lado interno do corpo eram suas pernas locomotoras e no lado
externo suas brânquias. Media entre 2,5 a 19 mm.
Naraoia


Naraoia é um gênero de pequeno a médio porte (cerca de 2 à 4,5 cm de comprimento), eram artrópodes marinhos dentro da família Naraoiidae, que viveram do início do Cambriano ao final do período siluriano. As espécies são caracterizadas por um grande sistema alimentar e antenas orientadas para os lados.

Naraoia morfologicamente é quase plano, e seu lado
superior (ou dorsal) do corpo consiste em um protetor de cabeça transversal ou
semi-circular não calcificado(encéfalo),e um escudo de cauda(pygidium) mais
longo que o encefalo, sem qualquer segmento corporal entre eles. O corpo é
estreitado na articulação entre o encéfalo e o piôndio. As antenas possuem
muitos segmentos são direcionadas para os lados. Não há olhos.
O intestino do Naraoia, possuia um diâmetro relativamente grande (14-18% da largura do corpo), e ao lado de quatro pares de grandes sacos digestivos. Possui apêndices com dois ramos em uma base comum, como os Misszhouia e trilobitas (ambos artrópodes cambrianos). Pelo menos os membros anteriores do tronco têm exópodes com lóbulos distais grandes em forma de remo e galhos laterais achatados curtos (cerdas) no eixo. O endópode (conhecido apenas no Naraoia compacta) é composto por seis podômeros.


Naraoia taijiangensis fóssil e em uma retocada ilustração da
publicação original de Walcott. Observe os apêndices à esquerda
Sedimentos presentes no intestino de
Naraoia sugerem que pode ter sido um alimentador de sedmentos,comendo grandes
quantidades de solo, como uma minhoca; Um sistema muito grande e complexo diverticular intestinal e um intestino com grande capacidade de retenção indicam
que a dieta de Naraoia Spinosa era baixa em nutrição. Por outro lado, a morfologia do sistema
digestivo também tem sido interpretada como representando um hábito predatório.
Os lobos distais grandes, em forma de remo e as cerdas lamelar curtas nos exópodes,
a implantação das antenas para o lado, e o ângulo do céfalo com o pilpídio de
até 90° com o qual muitos espécimes são encontrados, sugerem uma vida bentônica de de escavação do fundo sedimentar do mar.

Nenhum comentário:
Postar um comentário