O Apse dos Moluscos
Infauna Rasa
marinha.
A grande
variedade de formas deve-se, principalmente, aos diferentes hábitos alimentares
desenvolvidos. Os bivalves ( braquiópodes á maioria dos fósseis encontrados)
foram bem representados no Siluriano em registros fósseis, principalmente por
formas detritívoras e filtradoras formaram uma infauna rasa durante a
Era Paleozoica. Os cefalópodes, também dotados de rádula, adquiriram mandíbulas
fortes, tornando-se predadores e excelentes nadadores.
Os moluscos
fósseis representam organismos especialmente importantes do ponto de vista
paleoecológico, uma vez que a maioria dos grupos tem representantes vivos,
permitindo a utilização do princípio do atualismo na interpretação de sua
morfologia funcional. As diferentes formas e hábitos de vida dos moluscos
proporcionam dados importantes na reconstituição de ambientes antigos.
Os
tentaculitóides se tornaram proeminentes, e se espalharam pelos mares rasos
durante o período Siluriano, para aflorar por todos os mares tropicais e
subtropicais rasos da Terra.
Ecossistema marinho dos mares Silurianos
com grande maioria de Moluscos, Nautilóides e uma Comunidade diversa de
braquiópodes:
a)
Estrofomenídeo (Braquiópode Strophomenida)
b)
Leptaena (Braquiópode: Strophomenida)
c)
Onniella (Braquiópode: Orthida)
d)
Platystrophia (Braquiópode: Orthida)
e)
Brongniartella (Artropode: Trilobita)
f)
Calyptaulax (Artropode: Trilobita)
g)
Playtlichas (Artropode: Trilobita)
h)
bellerophontid (Molusco: (Monoplacophora)
i) orthoceratid (Molusco:Nautiloide)
j) Tentaculites (Cricoconarida)
Embora a maioria
das características morfológicas que definam as relações de parentesco entre as
diferentes classes de moluscos sejam parte moles de difícil preservação no registro,
a sobrevivência de diversos grupos ancestrais, a filogenia é bem resolvida
quando considerada a maioria dos grupos fósseis conhecidos.
É no período Siluriano que os Braquiópodes são os mais comuns organismos
com concha, compondo 80% das espécies totais como os Pentâmeras. Já os dos
Nautilóides estão no apogeu, e deixarão exemplares vivos até os tempos
modernos.
Espécime de braquiópode bem fóssil bem
conhecida, o Grammysia cingulata possuíam sua concha compostas originalmente de
carbonato de cálcio, que não foram preservadas. A forma dos moldes naturais (
dentro ) das conchas deixadas na rocha evidenciavam que eram bivalves. O
espécime foi encontrado em Cumbria( Inglaterra ) em camadas do período
siluriano entre 443 - 418 milhões de
anos atrás.
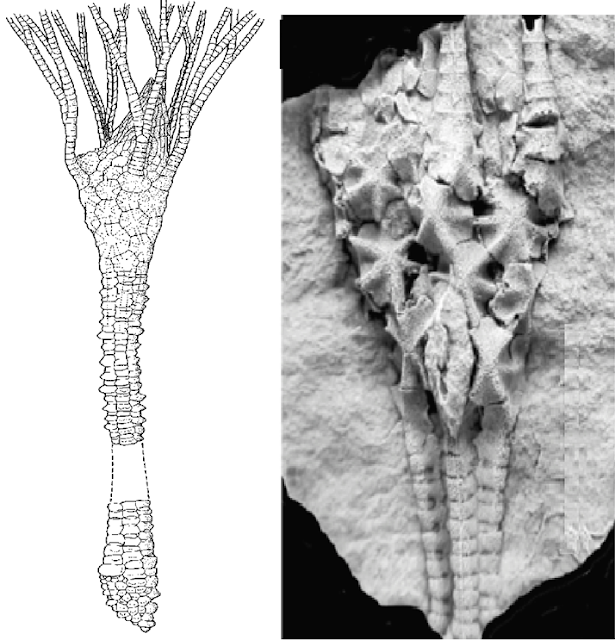
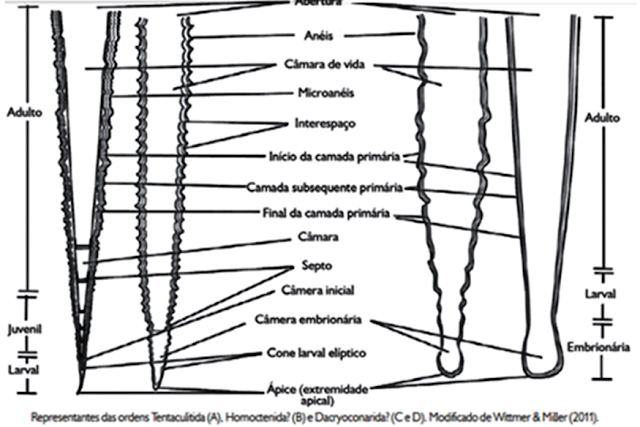
Tentaculitóides
Os tentaculitóides (do latim-tentáculo) foram
invertebrados marinhos reconhecidos pelo formato coniforme com uma seção
arredondada. As conchas eram de maioria carbonática e conchas calcítica; com
uma microestrutura semelhante a um braquiópodes.
As conchas eram lisas, com grandes
constrições ou cobertas com escultura na forma de anéis de vários tamanhos
aonde chegava de frações de milímetro a 30-50, mais frequentemente 70 mm, Na
América do Sul, o registro dos primeiros tentaculitóides ocorreu durante o início
do Siluriano, com o gênero Tentaculites.
Os Tentaculites apareceram no Siluriano, floresceram
no Devoniano Médio e desapareceram no final do Devoniano. Os fósseis se
observam desde os depósitos da extinção Ordovico-silúriana região equatorial
daquele período. Já no Siluriano a área de habitat fica mais concentrado na
região tropical e subtropical bem como no Devoniano Superior, quando dobra de
tamanho, as áreas ocupadas.
Vida nos mares dos Tentaculitóides
Lindstroemiella
eichwaldi
Fora um tentaculitóides incrustante da
Europa. O Lindstroemiella eichwaldi fora muito comum no Siluriano onde hoje é o
mar Báltico na Estônia. Ele possuía uma concha espiralada,
semelhante a um microconchido, com uma abertura orientada de forma variada,
sulcos transversais finos e afiados e sem uma morfologia semelhante a um cone
invertido. Mas levando em conta que tais tentaculitóides são
taxonomicamente e morfologicamente mais diversos do que se supunha anteriormente,
propomos incluí-los em uma ordem separada aa de Anticalyptraeida. Ao contrário
dos microconchídeos e cornulitídeos, os anticaliptraeídeos são o grupo menos
estudado de tentaculitóides e, portanto, pouco reconhecidos.
A era dos Nautiloídeos
Cena fictícia da Irradiação dos tipos de Nautilóides desde os de concha
reta até dos de concha enrolada.
Os cefalópodes
com concha (ectocócleos) externa (Nautiloidea) e interna (Endoceratoidea)
apareceram pela primeira vez no período Cambriano, cerca de 490 milhões de anos
atrás, e eram abundantes nos antigos oceanos da Terra. No geral destes animais,
eles possuíam flutuabilidade neutra e podiam sair do fundo e deslocar-se
lentamente pela coluna de água. A maioria das formas primitivas do Cambriano desapareceu,
mas uma única família, a dos Ellesmeroceratideos, sobreviveu até o fim do
Ordoviciano, onde acabou dando origem a todos os cefalópodes subsequentes.
Ellesmeroceratidae
Os cefalópodes de
concha reta a partir do período Ordoviciano, tiveram um imenso sucesso no
domínio dos mares onde se diversificaram em vários nichos como os Ortoconos.
Há denominação comum Ortocono é dada a praticamente todos os moluscos cefalópodes
nautilóides da era primária, que possuíam uma concha em forma de cone. Tais
moluscos assemelhavam-se muito às atuais lulas, com a diferença de possuir o
corpo no interior de uma concha.
Os cefalópodes que permaneceram em águas
rasas exibiram mudanças na forma da concha que melhoraram a capacidade de
natação e a capacidade de interagir com presas mais altas na coluna d'água em
comparação com suas contrapartes ancestrais. Muitos dos cefalópodes com conchas
longas e retas tinham a capacidade de flutuabilidade neutra em águas mais profundas
e um modo de vida no qual subiam e desciam diariamente na coluna d'água.
Tarphycerid Discoceras (Cephalopoda, Nautiloidea), do
Siluriano Inferior.
Mais para o final
do Ordoviciano começaram a ter sua concha enrolada, alterando assim o centro de
gravidade, o quê á tornou-se estruturalmente mais complexa. O pé evoluiu em uma
coroa de braços que permitia a captura de presas junto ao fundo. Deste modo
surgiram as subclasses Nautiloídea e Ammonoídea. Nautiloídeos tiveram seu
apogeu entre os períodos Ordoviciano e Siluriano (500-400 milhões de anos).
Possivelmente
oriundos do período Cambriano, no Ordoviciano Primitivo e Médio, os
nautiloídeos sofreram uma radiação evolutiva. Cerca de oito novas ordens
apareceram nessa época, cobrindo uma grande diversidade de tipos e estruturas
de conchas e estilos de vida ecológicos.
Ortoceras.
Os nautiloídeos
permaneceram no auge de sua gama de adaptações e variedade de formas entre os
períodos Ordoviciano, Siluriano e Devoniano, com várias formas de concha retas,
curvas e enroladas, todas coexistindo ao mesmo tempo. Várias das primeiras
ordens foram extintas nesse intervalo, mas outras ganharam destaque como a
Ortoceras.
Seção transversal
de um Ortoceras nautilóides do Siluro-Devoniano de Erfoud, Marrocos.
Os nautiloídeos
começaram a declinar a partir do Devoniano, Suas conchas ficaram cada vez mais
enroladas, enquanto o número e a variedade de espécies não nautiloídeos
continuaram a diminuir ao longo do Carbonífero e Permiano.
Estrutura Interna
das câmaras de um Nautiloídeos .

Evolução dos diferentes tipos de conchas em
cefalópodes Ectocócleos
A. concha ancestral; B. casca reta com endocones; C. concha com depósitos
anulares; D. concha com depósitos em câmaras; E. conchilla ascocerida; F.
concha
com fragmocone reduzido e câmara de sala com abertura voltada para baixo;
G-J. concha enrolada e com o fragmocone as partes moles; H. concha plana espiral
evoluída; I. Concha de Torticona; J. concha plano-espiral involuta.
O
fator Hidrostático
A compreensão das propriedades
hidrostáticas dos cefalópodes ectocócleos é essencial para o estudo da
morfologia funcional de suas conchas. As condições de flutuabilidade neutra, a
orientação estática do animal e sua estabilidade hidrostática são fatores-chave
na determinação de relações físicas e restrições nos hábitos de vida e
paleoecologia dos cefalópodes. Os tipos morfológicos foram escolhidos a partir
de diferentes ordens tradicionais, que incluem
os Cirtocones endogástricos (Ellesmerocerida
e Discosorida) e exogástricos.

Modelo
hidrostático de um ortocerídeo mesodômico (câmara do corpo ao comprimento total
= 33%). 1, Φgas distribuídos adapicamente. 2, Φgas distribuídos de forma
adorável. 3, Φgas distribuídos uniformemente dentro de cada câmera. 4, seção em
espiral. Verde = corpo mole, vermelho = gás cameral, azul = líquido cameral,
cinza = casca. A ponta da pirâmide vertical coincide com o centro de gravidade,
enquanto a ponta da pirâmide invertida coincide com o centro de flutuabilidade.
Φgas = porcentagem de fragmocone ocupado por gás cameral em flutuabilidade
neutra. St = índice de estabilidade.

Modelos
hidrostáticos do ellesmerocerid Plectronoceras. 1, Uma estimativa mínima
possível do comprimento da câmara corporal (40% do comprimento da câmara
corporal em relação ao comprimento total) estimada de Furnish e Glenister
(1964, p. K146, fig. 81, 1b). 2, Uma proporção de câmara corporal (35% do
comprimento da câmara corporal em relação ao comprimento total) quase
resultando em flutuabilidade neutra. 3, seção em espiral. Verde = corpo mole,
vermelho = gás cameral, cinza = casca. A ponta da pirâmide vertical coincide
com o centro de gravidade, enquanto a ponta da pirâmide invertida coincide com
o centro de flutuabilidade. Φ = porcentagem de fragmocone que precisaria ser
ocupada por gás cameral para atingir flutuabilidade neutra. St = índice de
estabilidade.
Todos os modelos cirtocônicos têm
estabilidade hidrostática semelhante à do Nautilus, e apenas o ellesmerocerid
tem flutuabilidade negativa. A hidrostática dos citocones de flutuação neutra
sugere que eles se comportaram como predadores, em vez de fracos nadadores
demersais ou nectobentônicos. Esse modo de vida coincide com uma distribuição
amplamente nerítica de indivíduos pertencentes a esses clados no Paleozoico. Em
contraste, os ortocones estudados eram capazes de flutuabilidade neutra, muito
mais estáveis do que o modelo
Náutilos, e todos tinham orientações
verticais syn vivo em um ambiente estático.

Modelo 3D
isométrico de Nautilus pompilius com corpo mole retraído (rente à abertura). 2,
modelo 3D isométrico de Nautilus pompilius com corpo mole estendido (modelo de
cabeça Náutilos fornecido por Peter Minister). 3, modelo 3D de Nautilus
pompilius criado a partir de um espécime digitalizado por TC (de Hoffmann et
al., 2014; Lemanis et al., 2015, 2016). Modelos hidrostáticos retraídos (4) e
estendidos (5) de corpo mole que têm gás e líquido de câmera distribuídos
uniformemente no fragmocone. 6, modelo hidrostático Nautilus pompilius
digitalizado por TC. Verde = corpo mole, vermelho = gás cameral, azul = líquido
cameral, cinza = casca. A ponta da pirâmide vertical coincide com o centro de
gravidade, enquanto a ponta da pirâmide invertida coincide com o centro de
flutuabilidade. Φgas = porcentagem de fragmocone ocupado por gás cameral em
flutuabilidade neutra. St = índice de estabilidade.
O declínio dos Nautilóides
Náutilos moderno.
As extinções
maciças no final do Permiano foram menos prejudiciais aos nautilóides do que a
outros taxa e alguns grupos sobreviveram até o início Mesozoico, incluindo
pseudo-torocerídeos, bactritídeos, nautilídeos e possivelmente ortocerídeos.
As últimas formas
de casca reta foram pensadas há muito tempo como desaparecidas no final do
Triássico, mas um possível ortocerídeo foi encontrado em rochas do Cretáceo.
Além dessa exceção, apenas uma única subordem nautilóide, a Nautilina, continuou
por todo o Mesozoico, onde coexistiram muito felizes com seus primos amonóides
mais especializados. A maioria dessas formas diferia apenas ligeiramente do
Náutilos moderno.
Eles tiveram um
breve ressurgimento no início Terciário ( talvez enchendo os nichos desocupados
pelos amonites na extinção K-T) e manteve uma distribuição mundial até o meio
do Cenozoico. Com o resfriamento global do Mioceno e Plioceno, sua distribuição
geográfica encolheu e esses animais resistentes e de vida longa caíram
novamente na diversidade. Hoje existem apenas seis espécies vivas, todas
pertencentes a dois gêneros, Náutilos ( o náutilos perolado ) e Allonautilos.




















































{kind=link}